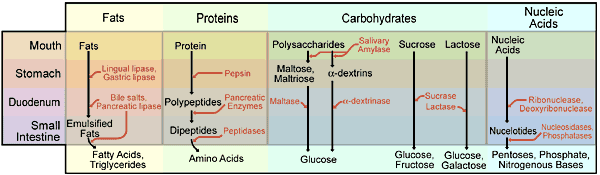
Table (280). The Digestion of the Principal Foods
Unit VI - Digestion
Chapter 20
Anatomy and Physiology of the Digestive System
1.The Function of the Digestive Process:
The foods we take in are assembled from components which are nothing like us either in their overall appearance or their intimate chemistry. In the digestive process, these foods are broken down mechanically and chemically until the breakdown products are small enough so that they can be absorbed. They are then re-assembled in the body in the way which is characteristic of the particular body. In this chapter we will be concerned primarily with the mechanisms involved in breaking down the foodstuffs so that they can be absorbed.
The mechanical part of the digestive process depends on the movements of the various parts of the digestive tract. These movements are mainly such as to reduce the size of particles within the tract. The smaller particles, mixed with the digestive secretions and under the influence of the digestive enzymes, undergo progressive chemical breakdown.
The chemical disintegration of foods is accomplished by specific enzymes. Usually foods and enzymes are suspended together in digestive secretions which are optimal for the action of the enzyme. Digestive enzymes, which are organic catalysts, accelerate food breakdown and are usually named after the substance broken down.
Before proceeding with this discussion, it may be helpful to identify the major foods and their components as well as the enzymes which break them down. This is done in the following table.
Digestive motility and secretion are largely controlled by extrinsic autonomic nerves, yet a surprising amount of digestion occurs in the absence of external nerves. Some is due to the formation of digestive hormones, some to the intrinsic nerves of the digestive tract.
Table (280). The Digestion of the Principal Foods
Note: Substances in the top row must be broken down by enzymes in order to be absorbed, while substances in the lowest row can be absorbed without difficulty. Enzymes are printed in red text, and note that enzymes which are introduced to the digestive process in the duodenum are originally secreted by the pancreas and liver. Ethyl alcohol, cholesterol, vitamins, and a small amount of triglyceride fats can be absorbed without enzymatic breakdown. This table is not complete.
2. The Structure of the Digestive Tract:
 |
 |
The digestive tract, or alimentary canal, runs from the mouth to the anus as a tube whose inside is a continuation of the skin. In a very real sense, then, undigested food is outside the body. Only after digestion, when it has crossed the wall of the digestive tract, is it inside the body. Many of the organs associated with the digestive tract are actually outpocketings of the tract. Thus, the liver and pancreas, major digestive organs, begin as simple glandular outpocketings of the first portion of the intestine. The simple outpockets themselves develop outpockets, which in turn develop outpockets, and so forth (Figure 281). Eventually, these complicated organs may be connected to the inside of the alimentary canal by a duct which is the remanent of the original outpocket, though the organ itself may be at some distance.
Figure 282 shows in generalized form the layers which are found in most of the tract. Most of these structures are also shown in the dissectograph. Note the dissectograph should be used to aid in understanding the relationships of the digestive organs to other organs.
3. Predigestion of Foods:
The digestive process actually begins before food is introduced into the alimentary canal, in some cases before it enters the home. For example, ground beef requires less mechanical breakdown in digestion than does the meat from which it is made. Similarly, many canned foods have been diced, which breaks them up mechanically, and precooked, which begins chemical breakdown. Another kind of chemical breakdown is accomplished by "hanging" meat before delivery. Dead tissues contain enzymes which begin the chemical breakdown of their proteins; hanging meat, in a sense, begins the digestive process before the meat is even delivered to the food market.
Cooking foods at home continues the process of breakdown, both mechanically and chemically. By slicing, prolonged heating, and using tenderizers, the fabric of the foodstuff is partially destroyed and its chemical decomposition enhanced.
By the time food is brought to the dining table, much of the digestive process which must be performed by animals has been begun. Even here, table silver is largely used to accomplish the further mechanical breakdown of the food.
 |
 |
4. Digestion in the Mouth:
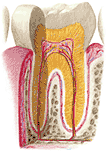
Even before food is put into the mouth, its smell may, through a conditioned reflex, produce salivary secretion. The saliva moistens and lubricates the food in the mouth and also contains a substance (salivary amylase) which begins the breakdown of food starches. The teeth continue the mechanical disintegration of the food begun in the predigestive processes, but they are not really required on a civilized diet. Normal teeth, shown in Figure 284, are covered with an extremely hard material (enamel), which can cut most foods and grind others. Within the enamel is a somewhat softer dentine, which in turn contains the pulp. The dental pulp contains the blood vessels and nerves which supply the tooth. Chewing the food cuts and grinds it while mixing it with saliva. Custom and habit govern the next step of the digestive process, which is swallowing.
5. Swallowing:
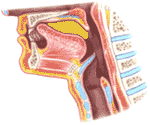
Swallowing occurs voluntarily, although the coordination of the muscles is very complex. The swallowing center of the medulla appears to control this regulation by way of the 9th, 10th, and 12th cranial nerves. The act begins with placement of the tip of the tongue against the hard palate; the sides of the tongue are also raised, but its mid-sagittal portion is depressed. Thus a chute is formed from which the only exit is the oropharynx.
The propulsion of food into the oropharynx is associated with elevation of its midportion, at first anteriorly, later posteriorly. The net result is that a bolus (ball) of food is forced back into the oropharynx. Potentially, the bolus entering the oropharynx might go upward into the nasopharynx or down into the airway. The first of these is prevented by muscular contractions in the tongue and soft palate which seal off the nasopharynx. The second is prevented by the upward movement of the laryngeal pharynx, which is sealed off by the elevation of the larynx to a position where it is covered by the epiglottis, which at the same time moves backwards. In addition, the laryngeal entry is closed by its own muscular effort. The bolus of food is thus propelled into the esophagus, which is the only avenue open to it. The nervous control is in the innermost portion of the brain, the medulla. The events of swallowing are animated in Figure 289.
 |
All of these events occur so effortlessly that they hardly ever reach the level of awareness. Yet their coordination may fail in the normal, and it commonly fails in old age and extreme weakness. When the nasopharynx is not well sealed off during swallowing, food may go into the nasopharynx and come out of the nostrils. If there is failure of epiglottic or glottic closure of the larynx, food may gain access to the airway. Usually, such an entry is rejected. The surface of the larynx is very sensitive to touch, and the response to failure to seal it off is usually a cough that expels the offending solid material.
This may not happen in a very ill or very weak person. In such a person food "swallowed the wrong way" may not be expelled at all but find its way into the airway and from there the lungs. Patchy infections of the lungs (bronchial pneumonia) may be the ultimate cause of death in such people.
6. The Esophagus:
The esophagus is altogether about 25 cm long. It has very important anatomical relationships. The heart lies just in front of its lower part, and the trachea is just in front of its upper part. The aorta arches around it and it perforates the diaphragm on its way to the stomach.
When the esophagus has been distended at its top by the entry of food, it tends to relax below and contract above the bolus. The effect of this, is of course, to move the bolus downward toward the stomach. This progression of a contraction wave, called peristalsis, is seen in almost every part of the gastro-intestinal tract.
Sometimes a piece of food which is large is taken into the esophagus. Peristalsis, which generates a weak force, cannot move it on. Here it induces pain, and the food is advanced only slowly by an esophageal force about which very little is presently known.
 |
7. The Stomach:
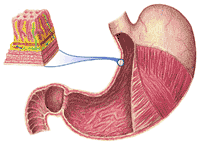
The stomach is shown in Figure 290 and can also be seen in its relationship to other structures in plates three and four of the dissectograph. It is a large organ which can hold a liter of fluid without trouble and can distend enough to hold four liters.
The relations to the surface of the body of the stomach are somewhat surprising to the layman. The stomach is an abdominal organ, yet most of it is behind the rib cage. Only the pyloric end of the stomach is uncovered by ribs.
Most of the stomach lies just under the diaphragm, which arches upward. Though the diaphragm separates the thoracic and abdominal cavities, its upward arching makes immediately sub-diaphragmatic structures appear superficially as if they were in the thorax. See Figure 291.
The wall of the stomach is made of three layers. The innermost contains secretory glands that produce the gastric juice. The middle layer is muscular, and the outermost is a thin tough sheet of fibrous tissue which is continuous with the lining of the abdominal organs, the peritoneum.
The stomach, like the esophagus, shows peristaltic waves that go from the cardiac to the pyloric end. The waves are incomplete; that is, they never result in complete closure of the stomach above and complete relaxation behind the wave. A peristaltic wave may reach a point in the stomach where a food particle is, but the food particle may move backwards as the wave advances since the cavity was never sealed.
There is a general tendency for food particles to move with the peristaltic wave, but as just noted, many particles, particularly large ones, may move backwards. Thus many hundreds of waves may pass over a food particle, each reducing its size a little until it is ready to move with the incomplete peristaltic wave. In effect the stomach "chews" the food.
In addition to this, the innermost lining of the stomach contains inumerable glands which form the enzyme pepsin and the hydrochloric acid which pepsin requires to act most effectively.
Pepsin begins the breakdown of protein molecules. They are not broken down by this enzyme to their simplest components (the amino acids) but to groups of amino acids (peptones).
The secretion of pepsin and acid by the stomach occurs in three phases. The first, like the salivary secretion which occurs when food is smelled, appears to be a conditioned reflex, depending on the sight or smell of food, even on the time of day. This is called the cephalic phase of gastric secretion. The entry of food into the antrum of the stomach now results in the release of a hormone, gastrin, into the circulation. This hormone, entering the blood draining the stomach, is in part returned to the stomach by way of arterial blood, and it further excites the stomach. This phase of gastric secretion, initiated and terminated in the stomach, is often referred to as the gastric phase. When the acid contents of the stomach enter the duodenum, a gastrin-like hormone may be produced. This phase is called the intestinal phase of gastric secretion.
The production of acid by the gastric glands is an extraordinary process which is not completely understood at present. Blood is a mildly alkaline fluid, while the gastric acid may be 0.1 N with respect to H+ ions. This represents a one-million fold increase in hydrogen ion concentration. It seems probable that the gastric glands accomplish the following reactions, one of which will be recognized as an absurd one by most chemistry students:
(1) H2O + CO2 <--> H2CO3
(2) H2CO3 + 2NaCl <--> NaHCO3 + 2HCl
Reaction (1) requires carbonic anhydrase, which is present in large quantities in the stomach. Reaction (2) can proceed toward the right only if one of its end products is removed. It is possible that this is just what happens. Minute amounts of HC1 formed in reaction (2) may be actively transported into the stomach. The NaHCO3 is carried away by the blood.
The manner in which the stomach empties was long a mystery. It seems clear now that peristaltic waves moving toward the antrum move the semi-liquid contents of the stomach toward the pylorus. For the most part, the pylorus is slightly open and little jets of gastric contents enter the next part of the digestive tract, the duodenum. How large such a jet will be will depend on the vigor and completeness of the peristaltic wave, the degree to which the pylorus is open, and the intrapyloric pressure. These factors are very closely integrated with each other by nervous and chemical mechanisms, and are not, at this time, fully understood.
8. Digestion in the Duodenum, the Liver, and Pancreas:
The duodenum, the first portion of the small intestine, plays a special part in the digestive process. It receives its name in an odd enough way; duodenum digitorum means twelve fingers breadths in Latin. The length of the duodenum is in fact 15 to 25 cm.
The importance of the duodenum depends on the fact that the two largest digestive glands empty here, the liver and pancreas. Furthermore, the duodenal mueosa is the site of formation of several hormones which regulate other digestive organs.
The anatomy of the duodenum is shown in Figure 292 and Plates 3 and 4 of the dissectograph. It consists of four parts. The first, connected to the pylorus, rises toward the right shoulder. The second proceeds directly downward and receives the duct of the pancreas and liver. The third part goes to the left. The fourth part rises toward the left shoulder until making a sudden ventral turn it connects to the next part of the small intestine, the jejunum. The whole of the duodenum fits around the head of the pancreas, which looks like a fist from the left pushing the duodenum to the right.
When gastric contents enter the duodenum, its lining layer, the mucosa, responds by the liberation of a number of digestive hormones. One of these, secretin, the first hormone discovered, (by Bayliss and Starling in 1902) enters the blood draining the duodenum. It will be recalled that the venous drainage of the digestive organs is by way of the hepatic portal vein. The secretin reaches the liver, passes through it into the hepatic vein and goes then to the right heart. A small but significant quantity now comes to the pancreas by way of its artery and excites the pancreas to secrete its digestive fluids, usually rather poor in enzymes.
Almost the same process results in the production of a hormone, pancreozymin, which stimulates enzyme production. However, the products of protein digestion rather than acid provoke the secretion of this hormone. Fats and acid entering the duodenum cause it to produce enterogastrone, which decreases gastric motility, and cholecystokinin, which causes the gall bladder to empty bile into the intestine. Other hormones produced in the duodenum will not be considered here.
This bewildering assortment of hormones makes sense when the sum total of their effects is considered. When stomach contents enter the duodenum, they call for digestive activity there. This is brought about by secretin, which produces the pancreatic juice and pancreozymin, which produces the pancreatic enzymes. Bile, necessary for fat digestion, is delivered by the gall bladder stimulated by pancreozymjin. Pancreatic digestion of fat is slow; the duodenum protects itself from being overwhelmed by producing enterogastrone, which adjusts the emptying of the stomach to the digestion of fats in the duodenum.
All of these events can take place entirely without nervous intervention, although there is undoubtedly some nervous control of the secretions of the pancreas, the emptying of the gall bladder, and the movements of the stomach, all of which are heavily innervated by both divisions of the autonomic nervous system. The function of nervous control of the digestive processes in the duodenum is not at all clear since these processes can take care of themselves. It may be that the innervation of these organs acts primarily to stop their activity when more pressing matters come to the attention of the organism, and to restart them when these matters have been disposed of. This is not, however, at all clear.
The pancreatic secretions are somewhat more alkaline than the blood and serve to neutralize the hydrochloric acid entering the duodenum from the stomach. This is necessary for the operation of the pancreatic enzymes, which do not act in acid media.
There are three important enzymes secreted in the pancreas:
(1) The carbohydrate-splitting enzyme pancreatic amylase.
(2) The fat-splitting enzyme pancreatic lipase.
(3) The protein-splitting enzyme trypsin. The pancreas actually secretes trypsinogen, a trypsin precursor which becomes active in the duodenum when acted on by an intestinal enzyme enterokinase.
These enzymes, mixed with the slurry of material leaving the stomach, continue the chemical breakdown of the principal foodstuffs.
The liver, which begins as an outpocketing of the gastro-intestinal tract, produces an important digestive secretion, the bile. It has, however, an impressive variety of non-digestive functions:
(1) Receiving the blood from almost the entire gastro-intestinal tract by way of the hepatic portal vein, the liver is ideally located to act as a storage organ for digested foodstuffs. In some cases the liver holds the absorbed materials, converting them to another form, and releases them slowly. This is well seen in the case of carbohydrates, which are absorbed as glucose, transported to the liver where they are converted to glycogen, which is released slowly as glucose. In other cases, the absorbed materials are converted by the liver into forms which can be delivered elsewhere. This function is best exemplified by considering the behavior of fat absorbed by the blood. Most of it comes to the liver as glycerol and fatty acids. These are reassembled (after some modifications) in the liver, combined loosely with proteins (lipoproteins) and sent to the fat depots. The amino acids from protein breakdown may undergo a variety of conversions--one of them is reassembly as proteins, which are released by the liver to the blood. Many vitamins, taken in the food or formed by bacteria in the intestine, are also stored in the liver.
(2) Apart from its storage function, the liver may be regarded as a metabolic factory where new compounds are made from old. Only a few examples will be used to illustrate this point. Carbohydrates may be converted into fat, and amino acids may be made from other amino acids or they may be converted into carbohydrates. Proteins may be made which contain different amino acids from those eaten by putting together newly formed amino acids. Chemical poisons may be combined with materials which make them harmless, and hormones present in excessive quantities can be destroyed.
(3) The digestive function of the liver is in the production of bile. Bile contains some of the materials of red blood cells which have come to the end of their life span, about 120 days. When these cells die, their hemoglobin is broken up into heme and the protein carrier. Heme, containing the iron of the hemoglobin molecule, is in part converted into bile pigments, which give the bile its greenish color.
From the standpoint of digestion, the bile salts are much more important. These are, essentially, soaps, that is to say, salts of fat-soluble acids.
Unlike the familiar household soaps, which are the sodium salts of fatty acids (or were before synthetic detergents took over), the bile salts are sodium and potassium salts of acids derived from oxidation of the cholesterol molecule. Their role is the emulsification of fat. This function depends on the solubility of the acid portion of the bile salt in both fat and water. In a sense, the bile acid, dissolving in both fat and water, brings substances dissolved in water into intimate contact with fat. See Figure 293. Thus, the pancreatic lipase, which is water soluble, is brought into contact with the fats in the duodenum, which are not, and their digestion can occur.
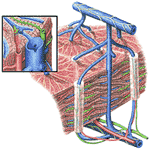 |
The anatomy of the liver is shown in Figures 294 and 295 and in Plates 3 and 4 of the Dissectograph. Figure 296 shows the microscopic structure of the liver in a three-dimensional reconstruction. Figure 292 shows the biliary system.
Delivery of bile to the duodenum at just the right time is normally accomplished by an elegant mechanism. The production of bile is a continuous process which occurs whether or not digestion is going on. It begins in the lobules of the liver which pass it into the bile capillaries. These fuse, eventually forming a large channel, the hepatic duct.
The hepatic duct branches into the common duct, which goes to the duodenum, and the cystic duct, which goes to the gall bladder. The duodenal opening is guarded by a circular muscle which is ordinarily tightly contracted. This muscle, the sphincter of Oddi, prevents passage of bile from the hepatic duct into the common duct. It therefore takes the alternative route, that is the cystic duct. Bile therefore normally flows from the liver to the gall bladder.
The gall bladder, in addition to serving as a receptacle for bile, acts to concentrate it. It takes up fluids through its walls leaving the bile salts behind. The solid content of bile on its way to the gall bladder is only 3%. This is increased four fold in the normal gall bladder through fluid reabsorption.
When fat enters the duodenum together with acid, cholecystokinin is released. This hormone causes relaxation of the sphincter of Oddi and contraction of the gall bladder. Concentrated bile enters the duodenum at the same time as pancreatic lipase, and both come in just after the stomach contents.
The hormonal mechanism for release of bile may or may not be supplemented by a nervous mechanism in health. The evidence that parasympathetic stimulation causes gall bladder contraction and relaxation of the sphincter of Oddi is convincing, though it is less clear that this actually occurs in the digestive process. It is, however, quite clear that parasympathetic drugs can be used to cause relaxation of the gall bladder and contraction of the sphincter even in the presence of cholecystokinin. This will be discussed further below in Part 13.
9. Digestion in the Small Intestine Past the Duodenum:
We know less about small intestinal digestion than digestion in any other part of the gastro-intestinal tract. Nevertheless, most of the work of digestion is done here.
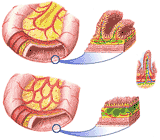 |
The small intestine is very long, about seven meters. The duodenum, which has already been discussed, is only 25 cm. long. The next 2.5 meters of small intestine is often found to be empty at autopsy, hence its name jejenum, empty. This section is not clearly separated from the last section, the ileum, which is about 3.75 meters in length. Sections of the jejunum and ileum are illustrated in Figure 298.
The structure of the mucosa of the small intestine favors absorption of digested materials. Finger-like projections, villi, extend into the cavity. These increase the surface available for absorption. They are richly supplied with blood vessels, all of which drain eventually into the hepatic portal vein and lymphatic capillaries, which drain into the thoracic duct by way of the cisternachyli. A large amount of it is carried by this system, whose capillaries, because of their milky appearance during fat digestion, are called the lacteals. These lymphatic channels are very heavily invested with lymph nodes, called the mesenteric nodes. These structures are illustrated in Figure 299.
Though it is so long, the small intestine is carried on a narrow sheet of tissue, the intestinal mesentery, the base of which is only 30 cm long. It supports all parts of the small intestine from behind. It is fan shaped, being much longer at its attachment to the small intestine than at its attachment to the back. See Figure 300.
All of the vessels and nerves of the small intestine lie in the mesentery. All foodstuffs after digestion travel in these vessels on their way to the liver via the portal vein. The nerves to the small intestine also travel to the liver through the mesentery.
The attachments of the small intestine are so loose that almost any type of motion can occur. It can sway on its mesentery; incomplete peristalsis can occur, as in the stomach; or it may shows segmental contractions (illustrated in Figure 301). Along with these movements, the small intestine secretes from its lining innumerable digestive enzymes which complete the disintegration and dissolution of its contents. Most of the soluble contents of the food which have not been absorbed before are absorbed here. By the end of the small intestine, there is practically no useful food left.
The absorption of foodstuffs from the small intestine occurs partly passively, but part of it is active. Proteins are absorbed only after they are broken down to amino acids; this absorption may be an active process. To a certain extent even the statement that proteins are absorbed only after breakdown may have exceptions. Small amounts of proteins, enough to produce food allergies, may be absorbed unchanged. Carbohydrates are absorbed in part by diffusion and in part by an active transport system involving phosphate.
Fats in the small intestine are absorbed by a process which is partly active. Some fats are split and reacted with phosphate to produce phospholipid, which serves to dissolve the rest; the remade and dissolved fats appear in the blood vessels and lymphatic vessels which course through the mesentery and eventually join the circulation.
Other nutrient substances, e.g. calcium, phosphate, iron (poorly), and vitamins are also absorbed in the small intestine. By the end of the small intestine, its contents are nearly devoid of nutritional materials. Water and salts are, however, still present.
10. Digestion in the Large Intestine:
The small intestine empties about four or five hours after a meal into the large intestine or colon. The anatomy of the colon illustrated in Figure 302. Its microscopic anatomy is shown in Figure 303. What little digestible material remains in the small intestine is digested and reabsorbed in the large intestine. Water is reabsorbed here, as are salts and the ordinary nutrients that have escaped absorption in the small intestine. The large intestine is heavily contaminated by bacteria, a fact which is distinct1y beneficial, for they produce trace materials which are useful to the organism, such as vitamin K.
The appendix, at the junction of small and large intestines, serves no known function. It can, however, cause serious trouble. When its drainage is blocked, its walls may distend and interfere with its own blood supply. Since it contains bacteria, it becomes infected very quickly. It may rupture, spilling its contents into the peritoneal space. This is a surgical emergency.
In ordinary circumstances the large intestine does not show the characteristic movements of the small intestine. Material entering it from the terminal ileum is stored in the ascending colon. Two or three times a day a peristaltic wave moves this material from the ascending colon to the sigmoid colon and rectum.
11. Defecation:
Defecation, which is largely under voluntary control, seems to be very much a matter of habit and custom. The idea of autointoxication by the bacteria of the large bowel is no longer accepted. In fact, the reverse seems to be true. Persons given large quantities of antibiotics which reach the colon may lose their colonic bacteria. As noted above, these bacteria produce vitamin K, and probably other important substances as well. Serious nutritional deficiencies may develop in such persons. They can be remedied by restoring the normal intestinal bacteria.
Feces, the material eventually eliminated from the bowel, is made up in part of indigestible food residues, the normal bacteria of the colon, and, to a very large extent, water. The remainder consists of insoluble and indigestible materials and has no nutritional value to animals, though it makes very good fertilizer for some plants.
12. The Hepatic Portal System:
Beginning with the lower end of the esophagus, and going on to the upper end of the rectum, the venous blood draining the bowel goes by way of the mesentery to the hepatic portal vein. This vein, whose course was indicated in Figure X, goes into the liver along with the hepatic artery. The common duct, carrying bile the other way, is associated with them.
Upon reaching the liver, the portal vein subdivides repeatedly. Eventually these blood vessels enter the liver lobules where they break up into sinusoids. These vessels are something like capillaries, but larger than most true capillaries. The flow of blood is from the periphery of the lobule, past the liver cells, to the vein at the center of the lobule (the central vein). The arrangement is shown in Figure 305.
Thus, all of the products of digestion which enter the blood stream are brought into close contact with the cells of the liver before being passed on to the heart. Only materials absorbed by the lacteals can bypass the liver, and these carry a very small fraction of the total of the materials absorbed.
There is, however, a significant exception to this statement. The veins which drain the upper part of the esophagus as well as those which drain the lower portion of the rectum do not empty into the portal system. They do, however, communicate with the veins which ordinarily drain the lower half of the esophagus and the upper part of the rectum. There is, therefore, an alternate pathway for blood ordinarily destined for the hepatic portal system. This pathway is extensively used in some diseases and may be used in some normal circumstances.
13. Disorders of the Digestive Tract:
It has already been indicated that the digestive organs are subject to autonomic control, although it is not clear that this control is actually used. It seems probable that this control is in reserve, although there are parts of the digestive tract in which it is of paramount importance. For example, the normal process of salivation appears to require parasympathetic stimulation of the salivary glands and also the absence of sympathetic stimulation. An extreme illustration of this has to do with the diagnosis of witchcraft. The suspected witch is asked to swallow dry bread. This is virtually impossible unless saliva is available as a lubricant. True witches, because of their guilt, have so much sympathetic activity that they cannot secrete saliva at all. Falsely accused witches terrified of the possibility that they may fail the test may have exactly the same problem. Though this would seem to make the test less useful than one would like, it nevertheless enjoyed great popularity in both Europe and America and has evidently proved successful in the eradication of witchcraft, since there are hardly any witches around anymore.
The evidence that emotion adversely influences the digestive processes below the esophagus is unsatisfying, which does not prevent the belief, part of most cultures, that proper digestion requires a proper nervous setting. Quarrels at meal time are considered likely to result in indigestion and are therefore avoided. Some restaurants soothe the nerves with innocuous music and subdued lighting, although the presumed advantages are probably lost as a result of the terror and dismay produced by examination of the prices on the menu or the final bill.
One of the most vexing popular ideas concerning indigestion which has successfully defied all evidence to the contrary for many years is that exercise, by diverting blood from the digestive organs to the muscles, predisposes to indigestion. While it is possible that exercise causes indigestion, it most certainly does not do so by reducing the blood supply to the digestive organs. Perhaps the most convincing demonstration ever offered of this was reported in 1967. Blood flow to one of the arteries of the intestine of a baboon was followed with an electromagnetic flow meter which broadcast its signals. The animals were permitted to resume normal life in the wild. There was no evidence in these animals that ordinary, even extraordinary, exertions would change intestinal blood flow. One animal was, in fact, killed by a carnivore during the experiment. Even during his death struggles, he showed no evidence of reduced intestinal blood flow.
Cavities occur in the teeth of most animals. Many animals die when they lose their teeth as a result of caries. Man, because of predigestion of food, easily survives dental losses.
The process of tooth decay appears to begin with the bacterial formation of acids which erode the enamel. When the enamel has been penetrated, bacteria can invade the dentine very rapidly. In many cases, the destruction of the dentine leaves the enamel a hard shell over an empty space; teeth so afflicted may suddenly crumble. When the infection comes near the pulp, bacterial products usually diffuse to the nerve, which they irritate, giving rise to toothache. Invasion of the pulp gives the causative organisms free access to the bone in which the affected tooth is lodged, and an abscess may result.
Enamel can be made extraordinarily resistant to decay by use of the fluoride ion, whether it is applied directly to the tooth from the outside or has been a part of the water supply. Fluoridation of water supplies has aroused much antipathy on the part of people who believe that the only legitimate business of government apart from warfare is reduction in federal, state, and local taxes, but there does not seem to be any medical or dental reason against it.
It has been noted that the esophagus lies behind the heart. Pain in the esophagus is often felt as pain in the area of the heart.
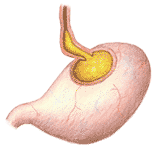 |
fig 306. The Esophagus in Heartburn |
There is no true valvular separation between the esophagus and the stomach. The angle at which the esophagus enters the stomach tends to prevent stomach contents from being forced into the esophagus during the contraction waves of the stomach, through a flap-valve like action. When the stomach is distended, particularly after dietary excesses, along with alcohol and tobacco (both of which cause excess stomach acid to form), the angle reduces, and the opening widens. Stomach contents can then be regurgitated into the esophagus (Figure 306). If the stomach contents are acid, there will be esophageal pain, burning in character, which is usually referred to as "heartburn". This is easily treated with antacid substances and easily prevented by avoiding volume excesses.
Peptic ulcers may occur as the result of self-digestion in the stomach, where they are called "gastric", or in the duodenum, where they are called "duodenal". Gastric ulcers are much less common. It cannot occur in the absence of gastric acid and pepsin, but its occurrence does not depend on over-secretion of these. In this form of peptic ulcer, the resistance of the cells to digestion is low. The stomach secretions do, in fact, digest the lining layer and proceed to the muscular layer, producing the ulcer "crater". In their course, they may digest the wall of an artery, leading to massive bleeding, a surgical emergency. They may ulcerate the heal over and over, leading to the formation of scar tissue, which impairs gastric motility and may cause obstruction. Occasionally, the ulcer proceeds through all the layers of the stomach, which then perforates. Stomach contents, including acid, are then disseminated through the abdominal cavity, where they cause great pain. This usually leads to shock, and the perforation must be repaired as an emergency measure.
The existence of a gastric (not duodenal) ulcer is occasionally the result of cancer, and this should always be considered when gastric ulcer has been diagnosed. In duodenal ulcers, gastric juice entering the duodenum may digest a portion of the duodenal lining; the consequences are in many ways the same as those in gastric ulcer.
The condition is common and annoying. Symptoms are quite regular. The entry of acid gastric juice into the duodenum causes pain (Approximately ten percent of all people have had it, and it is much more common in men than women). In gastric ulcer, as noted before, the secretion of gastric acid and enzymes tends to be normal or low. In duodenal ulcers the gastric secretion is usually high, but it may be normal in some cases.
The pain tends to occur one to two hours after eating when the acid stomach contents enter the duodenum. It is relieved fairly easily by antacid compounds, but these should not be considered curative. The pain tends to be located in the midline just under the breastbone and is often described as "boring" (as a drill) in character. The exact cause of the disease is unknown, but it seems quite clear that it will not occur unless acid and pepsin are both present in the gastric juices coming into the duodenum. Most treatments are directed toward reduction of gastric secretion or to neutralization of gastric acids.
A number of methods for reducing gastric secretion have been employed with more or less success. Alcohol, tobacco and coffee, all of which increase gastric secretion should be avoided by the patient with peptic ulcer. Some parasympatholytic drugs have been found quite useful. The use of bland diet and frequent small meals tends to neutralize the acids of the gastric juice before they reach the duodenum and is sometimes curative.
Surgical methods are used in many patients. A number of procedures are available. A few ulcers are associated with a pancreatic tumor which by some unknown mechanism causes the persistent secretion of acid and pepsin by the stomach. Removal of this tumor may be curative. Section of the vagal fibers to the stomach sometimes works, though some surgeons prefer to remove the greatest part of the secreting portion of the stomach.
It is popularly believed that there is a characteristic ulcer personality type. Most people tend to associate duodenal ulcers with the harassed executive personality. There does not seem to be any solid backing for this belief. Many attempts have been made to identify the ulcer personality. They have not been at all successful.
The normal flow of bile is from the liver to the gall bladder. When fat enters the duodenum, cholecystokinin is released. The gall bladder contracts and the sphincter of Oddi relaxes, and the bile flows into the intestine.
If there is a stone in the gall bladder, it may impair the normal outflow of bile from the gall bladder. This gives rise to intense pain (biliary colic) as the gall bladder contracts while the gall stone blocks the exit of bile from it. The cause of the contractions and pain has been described in Chapter 8.
Most physicians believe that gallstones which cause colic are an absolute indication for removal of the gall bladder. This writer tends to reserve judgement. The risks and discomforts of surgery are small but not negligible; the risks and discomforts of symptomatic gallstones may or may not be greater. No good statistics are available on this subject. (The student who wonders why may stop to reflect on the ethical problems of human experimentation. Can a doctor conscientiously withhold an enthusiastically recommended treatment from a suffering patient? Can he, on the other hand, recommend a treatment when he is not certain of the outcome of the disease or of the treatment ? These are old and vexing questions; their answers are not clear.)
Gall stones in the common duct, unlike stones in the cystic duct, threaten life, but rarely cause pain. The obstruction to the excretion of bile results in a significant failure in the absorption of fat and vitamin K, required for protein synthesis is not absorbed. The bile pigments, which are excreted, color the skin a brilliant yellow (jaundice). The accumulation of bile salts in the extra-cellular fluids may result in disastrous changes in their surface relationships to the cells.
Such stones should be removed. If the patient has sought medical attention because of jaundice, surgery should be deferred until an adequate level of prothrombin has been attained through injections of vitamin K. Oral administration of the vitamin is clearly useless since it will not be absorbed in the absence of bile in the duodenum.
Jaundice may also be produced through an acute inflammatory destruction of the liver tissue by a virus. The virus which brings this about may be transmitted from person to person by contact especially if that contact is made via a needle used for drug injections, and passed from person to person without proper sterilization. There are other ways in which this virus can be transmitted that usually involve the transmission of small amounts from material and result from carelessness with respect to elementary hygiene.
This is an extremely serious disease. It may produce great (sometimes fatal) destruction of the liver during the disease itself. This is rare but occurs. Much more frequently, the parts of the liver destroyed by the disease regenerate but in an anatomically peculiar way, and not completely. A few attacks may lead to a deformed, shrunken liver (Chronic hepatitis) which is not able to accomplish its normal functions. Such a liver, also called a cirrhotic liver, may partially obstruct the hepatic portal vein. The alimentary canal may drain through the alternate routes available to it: the anastomoses with systemic veins around the esophagus and rectum. These veins are not normally equipped to handle so much blood. They enlarge, sometimes to the point of rupture, leading (especially in the esophagus) to virtually uncontrollable hemorrhage. Although many cases of chronic hepatitis are of viral origin, others are due to malnutrition. In our society this usually results from alcoholism together with an inadequate diet. Opinions vary on the relative importance of the two, some maintaining that the alcoholic derives so much of his caloric intake from alcohol that he eats less of other foods. Others favor the idea that alcohol is the noxious agent, an idea not supported by experiments in dogs. One problem in evaluating the role of alcohol in chronic hepatitis is that alcoholics are notoriously untrustworthy, and the cirrhotic alcoholic may attribute his disease to anything that suits his fancy of the moment, readjusting his past history in such a way as to verify his ideas.
There is no curative treatment for established cirrhosis of the liver. The outlook for patients with the disease is very poor, 70% dying within five years. As noted above and in Chapter 14, the disease is often considered to be due to alcoholic excesses, and abstention from alcohol is vigorously recommended.
It was noted in Chapter 14 that surgical treatment does not produce particularly impressive results. This may be, in part, due to the fact that only those patients with the very worst outlook are considered for surgery.
Two types of surgical treatment have been recommended:
(1) connection of the portal vein to a systemic vein, such as the renal or splenic vein or even the inferior vena cava. This operation relieves the high venous pressure in the portal system. The liver, no longer in the main stream of the products of digestion, still receives them via the hepatic artery. The reduction of venous pressure in the portal system diminishes the chance or rupture of an esophageal varicose vein.
(2) It has been seriously suggested that the best treatment of a shrunken, damaged liver is the removal of most of it. The idea is that the liver will regenerate in an acceptable manner, which is quite true in the healthy liver. No serious evidence that the damaged liver will regenerate in a healthy way has been presented, and the operation is not often performed.
An odd and sad disease is fibrocystic disease. In this condition, seen most commonly in infants and children, the pancreas is virtually destroyed by mucus plugs in its ducts, and its digestive function is lost. The administration of pancreatic extracts is of value. Unfortunately, the lungs are usually affected as well. Alveolar air sacs, plugged at their bronchioles, become useless in the exchange of air and invite infection. Other alveolar air sacs enlarge and emphysema is a usual complication. This condition appears to be hereditary. At one time it was almost invariably fatal. At present, continuous, vigilant medical care extends life, though it is not curative.
These conditions may reflect the condition of the whole bowel or just the large intestine. In inflammatory disease of the small intestine, the colon may be swamped by non-absorbed material, and diarrhea results. Almost the same can be observed after taking magnesium sulfate, epsom salts; the small intestine cannot absorb more than a small quantity of this salt, and the unabsorbed part passes through the small intestine binding water. The large intestine cannot handle the extra load of fluid, and diarrhea results.
Constipation is more probably a condition of the large intestine and rectum. The normal urge to defecate occurs when the rectum is filled. If this urge is disregarded repeatedly, it may become a habit to disregard it. As noted above, defecation seems to be a matter of habit and custom rather than a regular phenomenon. Constipation may result from habitual neglect of feelings of fullness in the rectum; it may be due to an abnormal lack of the nerve cells which carry the propulsive wave through the colon. In either case, it does not seem to be particularly harmful. There is at least one case on record of constipation of one year's duration with no ill effects.
Continue to Chapter 21.