Unit V - Respiration
Chapter 17
Anatomy and Physiology of the Respiratory Passageways
1. The Function of Respiration:
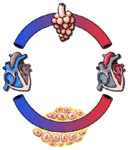
In one celled animals, the respiratory gases are all exchanged freely between the organism and its medium. Oxygen moves into such cells by diffusion, and carbon dioxide leaves in the same way. In larger animals, the cells are in contact with the outside medium--water in the case of fish, air in the case of mammals--only indirectly. Gas exchange is between the cells and the blood; in motion, the blood acts as a fluid bridge to the outside medium, where a second exchange occurs in the respiratory organs. The blood which leaves the tissues is usually poor in oxygen and high in carbon dioxide. That which returns from the respiratory organs, having been exposed to the outside medium, is higher in oxygen and lower in carbon dioxide.
|
These relationships are shown schematically in Figure 252. From this figure it should be apparent that normal respiration requires four things:
To illustrate how these functions must be integrated, some of the most common types of respiratory disturbance will be outlined here:
This chapter will be concerned with the anatomy of the respiratory organs and the mechanics of the process by which the outside air is brought near the blood in the lungs. In Chapter 18, some of the factors involved in the transport of the respiratory gases will be considered. Chapter l9 will be concerned with the mechanisms which adjust respiratory activity to the activity of the body.
2. The Upper Respiratory Passageways and Associated Structures:
In ordinary quiet respiration, the air which enters the lungs passes through the nose. The posterior part of the nasal cavity above the soft palate is called the nasopharynx. See Figure 253.
The nasopharynx, the oropharynx, and the laryngopharynx make up the pharynx, the area in which the respiratory and digestive pathways cross. This crossing is illustrated in Figure 254 and will be considered again in Chapter 19. The pharynx is lined by mucous epithelium. Its walls are made of voluntary muscles, which by appropriate contractions can seal off one or the other of the pathways.
During periods of increased activity, the mouth is also opened and breathing occurs through both the nasal pharynx and the oropharynx. Breathing through the oropharynx may substitute entirely for nasopharynged breathing when there is obstruction to the nasal passages, as in the common cold.
The air which enters the pharynx during inspiration passes by way of the larynx into the trachea, or windpipe, and from the trachea into the bronchi. The subsequent distribution of the bronchi will be considered in part 3.
The structures of the nasal cavity are shown in Figure 255. The entire nasopharynx is lined by a mucous membrane, as is the rest of the pharynx. The olfactory epithelium is at the top of the nasal cavity and not in the direct path of the air flow. Odors are more easily detected by "sniffing" than by ordinary breathing because "sniffed" air is brought into more direct contact with the olfactory mucosa than air which follows the respiratory pathway directly.
The inside lining of the trachea consists of a pseudo-stratified columnar epithielium. These cells, like cells in the nose, contain hair like processes, cilia, on the air side. The cilia appear to alternate between a slow bending downward and a whiplike action upward. The effect of this action is to expel upward solid or semisolid matter which has been inhaled or formed lower in the respiratory tree.
In mid-chest, the trachea divides
into two main bronchi, which are more or less similar to the trachea,
though somewhat smaller. Just above the level of division of the trachea is in
intimate relation to the arch of the aorta (Figure 263). The
bronchi are just behind the main pulmonary arteries. These relationships are
also shown in the Dissectograph.
3. The Lungs and Other Intrathoracic Structures:
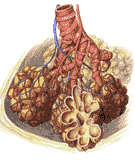 |
The two main bronchi divide into three branches on the right and two on the left. Each of the five bronchi so formed serves one lobe of the lung. Each such branch, a lobar bronchus, serves one lobe of the lung. As the bronchi continue to branch they gain in muscle while losing cartilage; eventually their walls are purely muscular, and they are called bronchioles. The terminal bronchioles terminate in alveolar ducts; these go into alveolar ducts which finally end as alveolar air sacs or alveoli of the lung. Here the outside air is brought into contact with the blood. Of course, it is not truly outside air; this may be appreciated when it is considered that the respiratory passageway is a two way street; the blood gases leave by the same route as the gases of the air enter, with inevitable admixture. The gas of the alveoli is called alveolar air. Its composition will be considered in the next chapter. The overall anatomy of the structures just described are illustrated in Figure 264.
The lungs lie in the thoracic cavity separated from each other by the mediastinum which contains non-respiratory intrathoracic structures. Its divisions are shown in Figure 265.
The most important area of the mediastinum is the middle mediastinum. Here, within the pericardium, are the heart, the root of the ascending aorta, the root of the pulmonary artery, and the atrial attachments of the superior vena cava, as well as the first division of the trachea. Lying on the pericardium are the two phrenic nerves on their way to the diaphragm. The superior mediastinum contains the arch of the aorta and its main branches, the veins which form the superior vena cava, the vagus nerves, the thoracic duct, trachea and esophagus. The posterior mediastinum contains the descending aorta, the vagus nerves and esophagus. All portions of the mediastinum contain lymph nodes and loose connective tissue.
In man, the mediastinum is relatively fixed and divides the chest into two hemithoraxes. This division is of great importance, for it enables either lung to function even when the other hemithorax is non-functional. Slight displacements, of the mediastinal structures to one side or the other are usually indicative of disease in one hemithorax.
The lungs themselves fill the two hemithoraxes in a rather peculiar way. Each lung is smaller (when removed) than its corresponding hemithorax. In the body, however, it fills the hemithorax almost completely. This results from the fact that there is no air in the thorax to exert a counter pressure against the air of the lungs, which is continuous with the outside air. The lung is pushed by the outside air against the thoracic wall. Except at its root, where there is an indirect connection to the thoracic wall, there is no anatomical connection at all. Thus each lung conforms to changes in the size of its hemithorax not by means of direct attachment but through the pressure of the air inside it uncompensated by air on the outside of the lung.
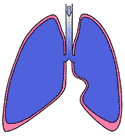
The pleura, a continuous membrane which lines the inside of the chest wall and also the outside of the lung, protects the lung structures from damage as they glide along the chest wall during respiration. The pleura also lines the outside of the mediastinum and the upper surface of the diaphragm. The parts of the pleura associated with the lung are called visceral pleura; the remainder is called parietal pleura. The parts of the latter associated with the chest wall are called costal, and the mediastinal and diaphragmatic pleura are related as their names indicate. These relationships are shown in Figure 266.
Figure 266 also shows that there is a space between the visceral and parietal layers of the pleura. This space, called the pleural sac, is small and filled with fluid in health. There are some areas where the space is a little larger. Here the lung cannot quite fill the cavity available for it, which is bent at an acute angle. The two most important such spaces are at either side, where the diaphragmatic pleura becomes the costal pleura. These spaces are called the costodiaphragmatic recesses. Another space corresponds to the sharp angle made between the pericardium on the left and the sternum.
The lungs within the visceral pleura are made of respiratory epithelium (the alveolar air sacs), and the passageways through which air must go to reach this epithelium (bronchial system). The blood supply of these two systems is different; the former receives its blood from the pulmonary artery and its branches, the latter from the bronchial arterial system. To a small extent these may intermingle in health, and their mixture may become of importance in disease. (See Figure 267).
The alveolar air sacs are in the most intimate contact with the pulmonary capillaries which surround them in the same way as a gauze network sac surrounds a flying balloon. The combined thickness of the alveolar membrane, the capillary wall, and the interstitium which separates them is about 0.1 micra, rather less than the thickness of most capillary endothelium (Figure 267). It will be recalled from Chapter 15 that the interstitium is kept nearly dry in the pulmonary circulation. The total respiratory surface is about 70-80 square meters.
 |
4. Mechanics of Breathing:
The combined acts of inspiration and expiration result in exchanges between the outside air and the air in the alveoli. As has been noted before, the respiratory passageways are a two way street, in which gases entering the alveolar air sacs from the arterial nd of the pulmonary capillaries are mixed with gases from the outside destined for the alveolar air sacs, where they will equilibrate with blood and leave the venular end of the alveolar capillaries. This arrangement, though it works, is less efficient than one in which air streams past the respiratory epithelium (see Figure 269). The inefficiency of the process is indicated by the fact that alveolar air contains only 2/3 as much oxygen as external air; its carbon dioxide content is 120 times as high as that of the outside air.
Some understanding of the reasons for this may be obtained by a simple calculation. The lung volume is about 5L; the amount breathed per breath is about 0.4L. Some 0.15 of the latter amount simply moves to and fro in the respiratory passageways, and the residue of some 0.25L is brought to the alveoli. Thus, 0.25L (exchanged gases) / 5L (total volume), or 5%, of the alveolar air is exchanged with each breath. 95% remains as it was.
This is such an inefficient process that one may wonder that it works at all. The fact that it does results from the fact that one breathes often, and that, when circumstances require, the volumes exchanged can be much larger. The role of frequency of breathing may be illustrated by the fact that at a normal breathing rate of 16 breaths / minute, although 95% of the alveolar air remain unchanged per breath, a little more than half the air (1 - 0.9516) is exchanged per minute. However, increases in rate of respiration, the depth remaining constant, produce more alveolar ventilation. The same changes just described may be brought about by 16 breaths taken in half a minute; in a minute the alveolar ventilation is enough to replace more than three quarters of the alveolar air. It is, however, much more effective to breathe deeply than to breathe rapidly. If, for example, the respiratory volume is doubled to 0.8 L, and 0.65 L reaches the alveoli per breath (the respiratory dead space remains constant in volume). Assuming 16 breaths / min, each breath replaces 0.65 L / 5 L, or 13%, of the alveolar air, 87% remaining as it was. In the course of a minute, 92% (1 - 0.8716) of the alveolar air is replaced. Both rate and depth increases are responsible for the increased alveolar ventilation of exercise.
How little each breath matters may be conveyed to the reader by a simple experiment. Ordinarily 4 breaths are taken during a quarter minute at rest. Very few persons experience any discomfort at all from a 15 second period of breath holding, which is the time required to drink from a glass of water (it will be seen later that there is no respiratory activity during swallowing). In the quarter minute of breath holding, the alveolar air, unchanged by respiratory activity increases its carbon dioxide concentration less than 20% while its oxygen also falls less than 20%.
The rate and depth of breathing are controlled by the central nervous system (See Chapter l9). The primary event in breathing is, however, a change in the size of the lung. Since the visceral pleura of the lung are in almost perfect contact with the parietal pleura, it follows that breathing will occur as the structures covered by the parietal pleura move. The structures principally involved are the chest walls and the diaphragm. Theoretically, the parietal pleura related to the pericardium are also involved, but since the pericardium is in almost the same position during systole and diastole of any of its chambers, respiratory function due to its movements can be neglected.
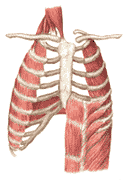 |
The movements of the diaphragm result in the most obvious volume changes in the lung. The diaphragm is dome shaped; when it contracts, flattening the dome, the space between the parietal pleura is increased. The visceral pleura, gliding freely along the parietal pleura, follow the increased volume, and the lung expands. Relaxation of the diaphragm results in a reduction in the volume of the lung. These volume changes can also occur as the result of changes in the configuration of the rib cage. This may be explaned by referring to Figure 270. This figure shows that in expiration, the sternum is sunk below the corresponding vertebra of the spinal column. The ribs angle downward from the spine to the sternum. If, by contraction of the external intercostal muscles, the ribs are brought upwards, there is an increase in the depth of the rib cage from front to back. This increases its volume and correspondingly the lungs expand. The internal intercostal muscles produce the opposite changes when they contract, increasing the downward slope of the ribs, the volume of the rib cage and the volume of the lung.
In normal persons, diaphragmatic and intercostal breathing go on together. In certain circumstances, one or the other may predominate, but which predominates appears to be a matter of individual training and circumstances existing elsewhere in the body.
Changes in lung volume involve all parts of the lung. For example, in inspiration, the bronchi and bronchioles dilate and lengthen, the amount of blood in the pulmonary venules and veins increases, and the alveolar air sacs increase in size and in number. These changes are all reversed on expiration. Some of them occur passively as the intrapleural pressure changes; others require active change (for example, inspiration is associated with a relaxation of the muscles of the bronchial system). One of the most interesting appears to occur passively, but it requires the presence of a chemical agent produced actively.
This has to do with the fact that the alveoli, like soap bubbles, tend to shrink more rapidly the smaller they get. A familiar illustration of this is seen when a soap bubble is blown from a pipe but remains attached. The bubble contracts slowly at first, then much more rapidly until the bubble disappears entirely, being replaced by a flat soap film over the orifice of the pipe. This phenomenon is due to the surface tension of the bubble which produces a much greater pressure in a small bubble than a large one. That this is so can be seen readily when two soap bubbles are blown from two pipes which are then connected at the mouth end. Invariably, the small bubble empties into the larger one.
 |
Applying this to the lung, it would seem that as the lung deflated, the large alveoli should grow larger while the small ones disappeared. This may actually happen in lung disease. In the healthy lung, however, it does not happen because all the alveoli are lined with a substance (surfactant) which reduces their surface tension. As an alveolus becomes smaller, its surface tension becomes smaller too, since its surfactant is distributed over a smaller area. As the lungs are distended, the larger alveoli--with the smallest surfactant concentration and therefore the highest surface tension--tend to resist further distension, but the smaller alveoli--high surfactant concentration, low surface tension--are easily inflated. Thus, the volume changes in the lung during inspiration and expiration are evenly distributed through the alveoli as well as the other structures of the lung. See also Figure 271.
5. Lung Volumes:
If one consciously suspends breathing activity in expiration, the lungs tend to assume a resting volume which is relatively constant for any one person. Normal inspiration increases the volume, the amount inspired being called the tidal air. It ordinarily amounts to about 400 ml.
Further inspiratory effort can result in further expansion of the lung. The extra volume which can be inspired is called the inspiratory reserve volume. This ordinarily amounts to about 3000 ml. The expiratory reserve volume is that volume which can be expired by effort after normal expiration, about 1200 ml.
After the most vigorous expiration, a volume of about 1200 ml remains in the lungs. This is called the residual volume.
Taken from the deepest possible
inspiration to the deepest possible expiration, the change in lung volume
including the inspiratory reserve, the tidal air, and the expiratory reserve is
called the vital capacity. This value does not include the residual
volume or the anatomical dead space (150 ml). In healthy young men it is about
4800 ml. These relationships are summarized in Figure 272.
6. The Work of Breathing:
The alternate increases and
decreases of the volume of the chest require that work be done. Some of this
work is used to overcome resistance to the movement of air in the respiratory
passageway, some is required to deform the lung itself, some to move the chest
wall, and some to move the organs which are displaced as the diaphragm moves.
The exact value is not known but it has been speculated
that the power involved in quiet breathing is about 5 watts. This is
almost 4 times the work of the whole heart. This value may rise almost
fifty times during maximum respiratory effort to about 250 watts. Man is
capable of using oxygen at a rate which corresponds to the development of 1000
watts for short periods; over longer periods (such as an hour), a power
consumption of 600 watts appears to be the limit of what man can use. However, respiratory effort is probably never more than 25%
efficient, which means that the lungs would have to use almost the full 1000
watts of maximum output in order to generate 250 watts. This leads to the rather
queer situation that the respiratory apparatus has greater capacity than can
ever be used except for very short periods, and even during such periods the
utilization of the full respiratory capacity to do work makes other work
impossible.
The answer to the apparent
paradox probably lies in the fact that the total respiratory capacity is never
used. Probably the limits of prolonged performance are set by the respiratory
apparatus, but at lower levels than those seen in respiratory "sprints". This is
probably not a significant factor in limiting the performance of normal men, but
it may become very important in disease.
7. Diseases of the Respiratory Passageways:
The list of respiratory diseases
is almost endless. A few illustrative examples will be chosen from each of the
above categories.
The common cold, or acute
coryza is, perhaps, the most common of all human illnesses. It is usually
caused by one of not less than 30 viruses. In itself, it is not a serious
disease, but it sometimes sets the stage for bacterial infections which may be
very dangerous.
The disease characteristically
involves the mucous membranes of the upper respiratory tract, including the
mucosa which lines the paranasal sinuses and the mucosa of the Eustachian tubes.
The larynx and sometimes the trachea may be involved.
The discomfort of the disease is
to a large extent due to the engorgement and inflammation of the mucosa. At
first this may be painful; later, when the mucosa begins to discharge watery and
mucous fluids, the principal discomfort results from the discharge itself. The
characteristic "stuffy nose," the sniffles, the headache (seen when the
paranasal sinuses are involved), and coughing or sneezing (as the discharges
irritate sensitive pharyngeal or laryngeal areas) all contribute to the misery
of the patient with a cold.
No effective method for
preventing colds has been devised as yet, nor is there any really effective
treatment. The patient can be made less unconfortable by the use of aspirin or
substances such as benzadrine taken from an inhaler. This tends to shrink the
swollen mucous membranes, but it may have undesirable side effects. Bed rest is
advisable, particularly when there is fever. Ordinarily, recovery is complete in
four to ten days.
Influenza is a much more
serious disease. The inflammation of the respiratory mucous membranes is
somewhat less than that seen in the cold. Constitutional symptons, such as
fever, weakness, muscular pains and loss of appetite are quite common. In many
cases, the inflammatory process spreads to the lower respiratory tract, leading
to an inflammation of some bronchioles and the alveolar air sacs which they
serve. This results in a patchy pneumonia (bronchial pneumonia). The outlook is
usually quite good, though in the influenza epidemic of 1918 (and to a much
smaller extent 1957) there were many deaths from pneumonia. Influenza vaccines
have proved very useful in the prevention of the disease although the immunity
they confer is not permanent. The treatment is symptomatic unless secondary
bacterial infections occur, in which case antibiotics should be used.
The lymphoid tissue which
surrounds the junction of the oropharynx and nasopharynx is ordinarily
protective. Bacteria from the nose and mouth are caught in this tissue. This
protective function may be lost, and the lymphoid tissue (tonsils and adnoids)
may become a source of difficulties when it is overwhelmed by infection. The
diseased tissue may then serve as a breeding ground for bacteria. Tonsillectomy
and adenoidectomy (removal of the tonsils and adenoid tissue) are often very
helpful.
By itself the common cold is
never disabling or fatal. It may, however, prepare the way for bacterial or
other virus infections which involve the entire respiratory system. Young
children are most often the victims. An apparently mild cold may explosively
become a laryngotracheabronchitis. The upper respiratory tract may be
blocked by mucosal secretions in the larynx; the lower respiratory tract may
also fill with mucous and large areas of lung may become useless in gas
exchange. Breathing is characteristically labored--this is the most obvious
symptom that something more than a cold is going on. This disease must be
treated promptly by a physician, for home remedies are virtually useless. Even
with the best medical care, many patients die.
Chronic bronchitis is a
chronic inflammation of the bronchial tree with persistent cough. It may be
carried by any of a number of organisms, by irritant substances in the inhaled
air (cigarette smoke and smog are common causes), by tuberculosis, or by other
internal causes. A persistent cough should always be considered as the sign of a
serious lung disease until proved otherwise.
Emphysema is an extremely
distressing disease for its victims. The alveolar air sacs are overdistended;
the useful area for gaseous exchange is reduced by obliteration of the
partitions between individual alveoli. The overdistended lungs also lose their
elastic properties. Ordinarily, when the chest is opened at autopsy, the lungs
collapse by virtue of their elasticity. In normal people in quiet breathing,
this tendency to collapse makes expiration quite automatic, though inspiration
requires some effort.
Foreign bodies occasionally
obstruct major bronchi or their subdivisions. This is most commonly seen in
infants and the elderly and infirmed. The portions of the lung served by the
obstructed bronchus are no longer ventilated and are usually poorly perfused.
They collapse as their gases are removed; while other parts the lung open up and
fill the space which was left by the collapsed parts. The collapsed portions of
the lung are said to be atelectatic; the expanded portions are
emphysematous. The treatment is removal of the foreign body, usually by
way of bronchoscope.
This picture is reversed in
emphysema. The lungs are fixed, as it were, in inspiration; and conscious effort
is required for expiration. In addition to this breathing abnormality, the
patient does not have enough respiratory surface to accomplish normal gas
exchange.
This disease is irreversible.
Insidious in its onset, it is usually relentless in its progression. It may be
of interest to note that there is more and more evidence to indicate that air
pollution plays a significant role in the development and course of the
condition. Just before this writing a group of physicians counseled against
living in Los Angeles, where emphysema is very troublesome, until atmospheric
pollution was reduced.
Pneumococcal pneumonia was
once very common. The causative agent (pneumococcus), makes pulmonary
capillaries permeable to proteins in the affected lobe or lobes. Pulmonary edema
results (See also Chapters 15 and 16). With adequate antibiotic treatment, survival
in this disease is close to 95%.
Pulmonary tuberculosis:
This disease is carried by the tubercule bacillus, an organism which can affect
any part of the body. The first inhalation of the bacteria usually occurs
without any symptoms at all. The bacteria are trapped, but usually not killed,
in lymph nodes at the root of the main bronchus of the affected lobe. Most
persons who have had one exposure to tubercle bacilli have no further trouble.
They develop a kind of hyperimmunity to the bacteria (Chapter 32) which is displayed when bacterial extracts
are injected in the skin, a tuberculis test. A positive tuberculis test,
indicating that hyperimmunity exists, is an almost certain sign that there has
been a previous exposure to the bacillus. The test is read as positive when the
injected area becomes reddened, hardened, and edematous two to three days after
injection.
A person who is hyperimmune
reacts very badly if reexposed to tubercle bacilli. Edema and swelling occur in
the lung just as in the skin. Further, the lung tissue (including bronchial
tissue and alveolar tissue) may be destroyed by the hyperimmune process. The
destroyed tissue is usually expelled in part by coughing. Some is reinhaled and
involves parts of the lung not involved before. The process may continue until
both lungs are massively involved and may be terminated by the death of the
patient. In the course of the disease, bronchitis, atelectasis, and tuberculous
pneumonias may all appear.
The decline in the death rate
from tuberculosis in the last half century has been nothing short of
spectacular. In 1900 the death rate from tuberculosis was 200 per 100, 000
population. By the l960s this figure had been reduced fifty fold. Part of
the decrease is due to improvement of the nutritional status of the population,
part is due to improved public health measures, and part to the advent of
specific agents which suppress the growth of the bacillus (streptomycin, para
amino salicylic acid, isoproniazid).
In some cases, patients with
tuberculous lung damage benefit from removal of a diseased lobe. Collapse
therapy (in which the affected lung is collapsed by the introduction of air
between the visceral and parietal pleura, by paralysis of one side of the
diaphragm, or by removal of several ribs on the affected side) is hardly ever
used, though it was once extremely popular in the treatment of tuberculosis.
Treatment by sun (heliotherapy) or by moving to a high, dry climate (climate
therapy) were both once in vogue, but both seem to be totally devoid of merit.
Inflammation of the pleura is
usually a sign of underlying disease of the lung. It is characterized by a
stabbing pain in the chest aggravated by respiration (pleurisy) The pain is due
to involvement of the parietal pleura and can usually be localized very
accurately. Such pleuritic pains should always be taken as evidence of serious
disease unless proved otherwise even if the pains disappear, as it is quite
likely to do if the involved pleural surfaces form fluid (pleurisy with
effusion).
Spontaneous pneumothorax
is a condition, by no means rare, in which the visceral pleura ruptures in an
overdistended area of lung. Air enters the intrapleural space and the lung
collapses a little. Increased respiratory effort increases the amount of
intrapleural air, bringing about further collapse. The process is usually halted
when the lung collapses enough to seal off the perforation.
The condition is rarely fatal,
and ordinarily no treatment is required other than bed rest until the ruptured
visceral pleura heals. The air in the chest is absorbed slowly and is usually
gone by the time the pleura has healed.
Perforating wounds of the
chest wall can lead to dramatically sudden collapse of the lung. The
respiratory distress experienced by the patient results in increasing the amount
of the pneumothorax. Sealing the wound, whether by a dressing impermeable to
air, or, in emergencies, by the application of the hand, may be life saving.
When the left heart fails and
enlarges, the pressure within its chambers rises. There is a corresponding rise
in the pressure in the pulmonary veins and venules which become swollen with
blood. Since the pulmonary venules make up part of the mass of the lung, their
swelling, which makes the lung heavier, makes breathing sufficiently difficult
to bring it to the level of awareness.
The consciousness of labored
breathing, called dyspnea, is one of the earliest signs of left heart
failure. It may only occur on exertion; but it may also occur from an act as
simple as lying down, which, by increasing the return of blood to the heart, may
overload the left ventricle.
In time, elevated pulmonary
venous pressure may be translated into increased pulmonary capillary pressure
and pulmonary edema. When pulmonary edema occurs the respiratory effectiveness
of the lungs is compromised. The arterial blood may be deficient in oxygen, so
that the heart suffers the further disadvantage of working in an unfavorable
(negatively inotropic) chemical environment and dilates further, thus
increasing venous pressure. The steps in this cycle usually follow each other
inexorably; however, the cycle may be terminated by right heart failure (the
right heart becoming unable to pump against the increased pulmonary capillary
pressure). Vigorous and determined treatment may also interrupt the cycle.
Bleeding diminishing the work of the heart, digitalis, improving the chemical
status of the heart muscle, and the lessening of venous return by the
application of loose tourniquets to arms and legs may be combined with oxygen
administration, which gets more oxygen into the blood than is supplied by
ordinary air in the presence of pulmonary edema.
Almost all of the above changes
can occur when the atrioventricular valves on the left side of the heart are
stenotic (mitral stenosis) even though the left ventricle muscle is normal. This
condition can be treated best by valve replacement; in some cases the stenotic
valve can be forced open by pushing a finger through it after opening the chest.
The finger need not enter the heart, but rather is pushed along with atrial
tissue through the stenotic valve. This procedure, called closed mitral
commissurotomy, is almost without risk in skilled hands and results in
improvement in 80-90% of all cases. See also Chapter
16.
In Part 6 it
was shown that quiet breathing in health required 5 watts of power. It was also
noted that the respiratory muscles are at most 25% efficient. It will be shown
in Chapter X than the energy available to a
resting man is equivalent to 75-80 watts. Thus the respiratory muscles (which
develop 5 watts at 25% efficiency) use no less than 20 watts at rest, about one
quarter of the power available to the body.
When part of the respiratory
passageway is obstructed, the power demanded by the increased respiratory effort
may increase faster than the oxygen supply made available through that effort.
When this occurs, all the available oxygen may be spent in the respiratory
endeavor while other organs suffer.
The extent to which respiratory
disease increases respiratory effort and exaggerates respiratory need is not
known with any degree of certainty. It does, however, seem quite probable that
minor disturbances in respiratory function may lead to major respiratory
distress. This area deserves further study.
Continue to Chapter 18.
a. Upper Respiratory Tract Diseases:
b. Diseases of the Lower Respiratory Tract and Lungs:
c. Diseases of the Pleura and Chest Wall:
d. The lung in heart failure:
e. The work of breathing in disease: